
میانهزیستی
| زمانبندی زمینشناسی | |||
| پیدازیستی (فانروزوئیک) | نوزیستی (سنوزوئیک) | کواترنری | |
| نئوژن | |||
| پالئوژن | |||
| میانهزیستی (مزوزوئیک) | کرتاسه | ||
| ژوراسیک | |||
| تریاس | |||
| دیرینهزیستی (پالئوزوئیک) | پرمین | ||
| کربونیفر | |||
| دوونین | |||
| سیلورین | |||
| اردویسین | |||
| کامبرین | |||
| نهانزیستی (پرکامبرین) | پیشینزیستی (پروتروزوئیک) | ||
| نخستزیستی (آرکئن) | |||
| پیشازیستی (هادئن) | |||
میانهزیستی یا مزوزوئیک[۱] (به انگلیسی: Mesozoic) یکی از سه دوران در ابردوران پیدازیستی است که مابین دورانهای دیرینهزیستی و نوزیستی قرار دارد و از حدود ۲۴۵ تا ۶۵ میلیون سال پیش را شامل میشود.
این دوران ۱۶۰ میلیون سال به طول انجامید و شامل سه دوره به نامهای تریاس، ژوراسیک و کرتاسه بود.
جانورانی مثل شاخقوچی (آمونیت) و دایناسور از جاندارانی بودند که در این دوره زیسته و منقرض شدهاند. خزندگان درشتجثهٔ میانهزیستی را دایناسور مینامند. آب و هوای این دوره گرم و استوایی بوده و نسبت به دوره ما بسیار باثبات بودهاست. خشکیهای زمین در این دوره شامل قاره لوراسیا و قاره گندوانا بود؛ که لوراسیا که شامل قاره آسیا و اروپا و آمریکای شمالی و گرینلند بود در دوره کرتاسه آغاز به تجزیه گرفتهاست و از این طریق اقیانوس اطلس به وجود آمده؛ و قاره گندوانا در دوره تریاس تجزیه شد.
دوره میانهزیستی با سلطه بازدانگان و خزندگانی مانند دایناسورها؛ آب و هوای گرم گلخانهای؛ و تجزیه تکتونیکی ابرقاره پانگهآ مشخص میشود.
میانهزیستی بهدنبال انقراض انبوه پرمین-تریاس آغاز شد (بزرگترین مورد مستند شده انقراض جمعی در تاریخ زمین)، و با انقراض انبوه کرتاسه-پالئوژن پایان یافت. این مورد دوم نیز انقراض دیگری بود که قربانیان آن شامل دایناسورهای غیرپرنده، پتروسورها، موساسوریان و پلسیوسورها بود. میانهزیستی دورهای از فعالیت قابل توجه زمینساختی، اقلیمی و فرگشتی بود. این دوره شاهد تکهتکه شدن تدریجی ابرقاره پانژهآ به تودههای خشکی جداگانهای بود که در دوره بعدی به موقعیت فعلی خود منتقل شدند. آب و هوای میانهزیستی متنوع بود و بین دورههای گرمایش و سرمایش متناوب بود. با این حال، بهطور کلی زمین گرمتر از امروز بود. دایناسورها برای اولین بار در اواسط دوره تریاس ظاهر شدند و در اواخر تریاس یا اوایل ژوراسیک به مهرهداران زمینی غالب تبدیل شدند و این موقعیت را برای حدود ۱۵۰ یا ۱۳۵ میلیون سال تا زمان نابودی خود در پایان دوره کرتاسه حفظ کردند. پرندگان ابتدایی در دوره ژوراسیک ظاهر شدند که از شاخهای از دایناسورهای دَدپا تکامل یافته بودند، سپس پرندگان حقیقی بیدندان در کرتاسه ظاهر شدند. اولین پستانداران نیز در طول میانهزیستی ظاهر شدند، اما تا دوره نوزیستی کوچکجثه باقی ماندند (کمتر از ۱۵ کیلوگرم). گیاهان گلدار در اوایل کرتاسه ظاهر شدند و به سرعت در سراسر پایان دوره گونهگون شده و جایگزین مخروطیان و سایر بازدانگان، مانند کهندارسانان، نخلیسرخسسانان و بنتیتسانان[۲] به عنوان گروه غالب گیاهان شدند.
نامگذاری
[ویرایش]عبارت «عصر خزندگان» در قرن نوزدهم توسط دیرینشناس، گیدئون مانتل، معرفی شد. او این دوران را تحت سلطه خزندگان دوکمانی مانند ایگوآنودون، مگالوسور، پلسیوسوروس و بالانگشتیها میدانست.
نام کنونی (مزوزوئیک) در سال ۱۸۴۰ توسط زمینشناس بریتانیایی، جان فیلیپس (۱۸۷۴–۱۸۰۰) پیشنهاد شد. «مزوزوئیک» در لغت به معنای «زندگی میانی» است که از پیشوند یونانی باستان -meso (به معنای «بین») و zōon (به معنای «حیوان، موجود زنده») مشتق شدهاست.[۳][۴] بدین ترتیب، مزوزوئیک با دورههای سنوزوئیک («زندگی جدید») و پالئوزوئیک («زندگی قدیمی») و همچنین ابردوران پروتوزوئیک («زندگی اولیه») قابل مقایسه است.
دوره میانهزیستی در ابتدا به عنوان دوره «ثانویه» توصیف شد؛ یعنی پس از دوره «اولیه» (پالئوزوئیک) و قبل از دوره ترشیاری (دوره سوم).[۵]
دورههای زمینشناسی
[ویرایش]| دوره | دور | عصر | سالها (به میلیون) |
|---|---|---|---|
| کرتاسه | پسین | ماستریختین | ۶۶٬۰ - ۷۲٬۱ |
| کامپانین | ۷۲٬۱ - ۸۳٬۶ | ||
| سانتونین | ۸۳٬۶ - ۸۶٬۳ | ||
| کنیاسین | ۸۶٬۳ - ۸۹٬۸ | ||
| تورونین | ۸۹٬۸ - ۹۳٬۹ | ||
| سنومانین | ۹۳٬۹ - ۱۰۰٬۵ | ||
| پیشین | آلبین | ۱۰۰٬۵ - ۱۱۳٬۰ | |
| آپشین | ۱۱۳٬۰ - ۱۲۵٬۰ | ||
| بارمین | ۱۲۵٬۰ - ۱۲۹٬۴ | ||
| هاتریوین | ۱۲۹٬۴ - ۱۳۲٬۹ | ||
| والانجینین | ۱۳۲٬۹ - ۱۳۹٬۸ | ||
| بریاسین | ۱۳۹٬۸ - ۱۴۵٬۰ | ||
| ژوراسیک | پسین (مالم) |
تیتونین | ۱۴۵٬۰ - ۱۵۲٬۱ |
| کیمریدگین | ۱۵۲٬۱ - ۱۵۷٬۳ | ||
| آکسفوردین | ۱۵۷٬۳ - ۱۶۳٬۵ | ||
| میانه (داگر) |
کالووین | ۱۶۳٬۵ - ۱۶۶٬۱ | |
| بتونین | ۱۶۶٬۱ - ۱۶۸٬۳ | ||
| باجوسین | ۱۶۸٬۳ - ۱۷۰٬۳ | ||
| آلنین | ۱۷۰٬۳ - ۱۷۴٬۱ | ||
| پیشین (لیاس) |
توآرسین | ۱۷۴٬۱ - ۱۸۲٬۷ | |
| پلینسباکین | ۱۸۲٬۷ - ۱۹۰٬۸ | ||
| ساینمورین | ۱۹۰٬۸ - ۱۹۹٬۳ | ||
| هتانگین | ۱۹۹٬۳ - ۲۰۱٬۳ | ||
| تریاس | پسین | رتین | ۲۰۱٬۳ - ۲۰۸٬۵ |
| نورین | ۲۰۸٬۵ - ۲۲۸ | ||
| کارنین | ۲۲۸ - ۲۳۵ | ||
| میانه | لادینین | ۲۳۵ - ۲۴۲ | |
| آنیسین | ۲۴۲ - ۲۴۷٬۲ | ||
| پیشین | اولنکین | ۲۴۷٬۲ - ۲۵۱٬۲ | |
| ایندون | ۲۵۱٬۲ - ۲۵۲٬۲ |
دورهٔ مزوزوئیک یا میانهزیستی به دنبال دیرینهزیستی، تقریباً به مدت ۱۸۶ میلیون سال ادامه داشت؛ از ۲۵۱٫۹۰۲ تا ۶۶ میلیون سال پیش، آغاز عصر نوزیستی. این بازه زمانی به سه دوره زمینشناسی تقسیم میشود. از قدیمیترین تا جدیدترین آنها عبارتند از:
- تریاس (۲۵۱٫۹۰۲ تا ۲۰۱٫۴ میلیون سال پیش)
- ژوراسیک (۲۰۱٫۴ تا ۱۴۵ میلیون سال پیش)
- کرتاسه (۱۴۵ تا ۶۶ میلیون سال پیش)
مرزِ پایین میانهزیستی توسط رویداد انقراض پرمین – تریاس تعیین میشود که طی آن، تخمین زدهشدهاست که تا ۹۰ الی ۹۶ درصد از گونههای دریایی متحمل انقراض شدند.[۶] گرچه با برخی برآوردهای دیرینشناسان که این را تا ۸۱٪ اعلام میکنند، بر سر این اعداد و ارقام سؤال مطرح شدهاست.[۷] همچنین به دلیل اینکه یکی از بزرگترین انقراضهای جمعی در تاریخ زمین محسوب میشود، از آن به عنوان «مرگ بزرگ» یاد میشود.
مرز بالایی مزوزوئیک در رویداد انقراض کرتاسه-پالئوژن (یا انقراض (K-Pg) تعیین میشود[۸] که ممکن است در اثر برخورد سیارکی بهوجود آمده باشد که دهانه چیکشلوب را در شبهجزیره یوکاتان پدیدآورده است. باور بر این است که به اواخر دوره کرتاسه، فورانهای بزرگ آتشفشانی نیز به رویداد انقراض کرتاسه-پالئوژن کمک کردند.[۹] تقریباً ۵۰٪ از همه سردهها منقرض شدند، شامل تمام دایناسورهای غیرپرنده.
تریاس
[ویرایش]دوره تریاس تقریباً از ۲۵۲ میلیون تا ۲۰۱ میلیون سال پیش را در بر میگیرد و قبل از دوره ژوراسیک قرار دارد. این دوره بین رویداد انقراض پرمین–تریاس و رویداد انقراض تریاس–ژوراسیک، دو مورد از «پنج مورد بزرگ» محصور شدهاست و به سه دوره اصلی - تریاس اولیه، میانی و پسین - تقسیم میشود.[۱۰]
تریاس اولیه، حدود ۲۵۲ تا ۲۴۷ میلیون سال پیش، تحت نفوذ بیابانهایی در فضای داخلی ابرقاره پانگهآ بود. زمین به تازگی شاهد یک مرگ گسترده بود که ۹۵ درصد تمام زیستمندان در آن ناپدید شدند و رایجترین مهرهداران روی زمین بیلخزندهها، مازدندانان و هوپارکریها همراه با بسیاری از موجودات دیگری بودند که از انقراض پرمین جان سالم به در بردند. بریدهمهرگان در اوایل تریاس به حداکثر تنوع خود رسیدند.[۱۱]

تریاس میانی از ۲۴۷ تا ۲۳۷ میلیون سال پیش، شامل فرایند ابتدایی تجزیه پانگهآ و شکلگیری اقیانوس تتیس بود. اکوسیستمها از انقراض پرمین احیا شده بودند. جلبکها، اسفنجها، مرجانها و سختپوستان همگی بهبود یافته بودند و خزندگان دریایی جدیدی - مانند ایکتیوسورها و کاذبخزندهسانان - تکامل یافتند. در خشکی، جنگلهای کاج شکوفا شدند، همانطور که گروههایی از حشرات شامل پشهها و مگس میوه، گسترش یافتند. خزندگان رو به بزرگ شدن گذاشتند و نخستین تمساحها و دایناسورها تکامل یافتند. به ترتیب رقابتهایی با دوزیستان بزرگ (که قبلاً بر دنیای آب شیرین حکومت میکردند) و خزندگان پستاندار در خشکی آغاز شد.[۱۲]
پس از رونق دوره تریاس میانی، دوره تریاس پسین از ۲۳۷ تا ۲۰۱ میلیون سال پیش، گرمای مکرر و بارش متوسط (۲۵–۵۰ سانتیمتر در سال) را دربرداشت. گرمای تازه منجر به شکوفایی ناگهانی و گسترده تکامل دایناسورها بر زمین شد، زیرا قارهها شروع به جدا شدن از یکدیگر کردند (نیاساسور از ۲۴۳ تا ۲۱۰ میلیون سال پیش - تقریباً ۲۳۵ – ۳۰ میلیون سال - برخی جداییها شکل گرفت که موجب پیدایش خزندهپاریختان، ددپایان و خزندهریختان شد) این شرایط همچنین موجب تکامل و ظهور گونههای اولیه پتروسورها شد. در طول تریاس پسین، بعضی از سگدندانهای (پیشرفته) موجب پیدایش پستاندارسانان اولیه شدند. اما همه این تغییرات اقلیمی منتج به مرگ و میر بزرگی معروف به رویداد انقراض تریاس-ژوراسیک شد که بسیاری از شاهخزندگان (به جز پتروسورها، دایناسورها و تمساحریختان) بیشترین همکمانان، و تقریباً تمامی دوزیستان بزرگ منقرض شدند. به علاوه ۳۴٪ زیستمندان دریایی در طی چهارمین رویداد انقراض جمعی منقرض شدند. دلایل این انقراض همچنان با ابهام روبروست؛ فورانهای سیل بازالتی در استان ماگمایی مرکز آتلانتیک یکی از دلایل احتمالی مطرح شدهاست.[۱۳][۱۴]
ژوراسیک
[ویرایش]
دوره ژوراسیک بین بازه زمانی ۲۰۰ تا ۱۴۵ میلیون سال پیش واقع شده و سه دورهٔ اصلی یعنی ژوراسیک اولیه، میانی و پسین را تشکیل میدهد.[۱۵]
ژوراسیک اولیه از ۲۰۰ تا ۱۷۵ میلیون سال پیش را در بر میگیرد.[۱۵] آب و هوای گرمسیری آن، بسیار مرطوبتر از آب و هوای تریاس بود؛ این وضعیت پیامد ظهور دریاهای پُر وسعت بین تودههای خشکی محسوب میشود. در اقیانوسها پلسیوسورها، ایکتیوسورها و شاخقوچیها فراوان بودند. بر روی خشکی، دایناسورها و دیگر شاهخزندگان به عنوان نژاد غالب حکومت میکردند و ددپایانی مثل دیلوفوسور در بالای زنجیره غذایی بودند. نخستین تمساحهای واقعی تکامل یافتند و دوزیستان بزرگ را به سمت انقراض هل دادند. نتیجه آن شد که شاهخزندگان در سراسر گیتی قدرت گرفتند. در همین حال و از سوی دیگر اولین پستانداران واقعی تکامل یافتند، گرچه بهمراتب کوچکتر بودند ولی بهطرز عجیبی همهجا پراکنده شدند. برای نمونه «بیدستردنب» گونهای متعلق به ژوراسیک، سازگاریهایی برای شنا، کندن و گرفتن ماهی کسب کرده بود. پستاندار «فروتاکاونده» از اواخر دوره ژوراسیک که تقریباً ۱۵۰ میلیون سال پیش وجود داشت، تقریباً اندازه یک سنجاب را داشت و دندانها، اندامهای جلویی و مهرههای ستون فقرات آن نشان میدهد که لانه حشرات اجتماعی (احتمالاً موریانهها، زیرا مورچهها هنوز پدیدار نشده بودند) را میکند؛ «بالداردد» نوعی دیگر از پستانداران بود که تواناییِ خزیدن برای مسافتهای کوتاه را داشت؛ درست شبیه سنجابهای پرنده کنونی. همچنین از نخستین نمونه «چندتکمیزهایان» مانند «روگوسودون» تکامل یافتند.
ژوراسیک میانی بین ۱۷۵ تا ۱۶۳ میلیون سال پیش قرار میگیرد. در طول این دوره دایناسورها شکوفا شدند و گلههای غولپیکری از خزندهپاریختان مثل برکیوسور و دوتیرکی چراگاههای سرخسی را شخم میکردند و تحت تعقیب شکارچیان تازهفرارسیده و متعددی، از جمله آلوسورها بودند. جنگلهای مخروطیان بخش بزرگی از جنگلهای سراسر زمین را اشغال کرده بودند. درون اقیانوسها، پلسیوسورها کاملاً معمولی بودند و ایکتیوسورها فراوانی داشتند. اوج شکوفایی خزندگان در همین دوره روی داد.[۱۶]

ژوراسیک پسین از ۱۶۳ تا ۱۴۵ میلیون سال پیش طول میکشد. در این دوره، اولین پرندهبالان از گونه تکاملیافته دایناسورهای تُهیدُمخزنده کوچک - مثل آرکئوپتریکس (دیرینهمرغ) - پدیدار شدند. افزایش سطح آب دریاها راه دریی آتلانتیک را گشود، آبراههایی که تا امروز پیوسته بزرگتر شده و درحال گسترش است. جدایی بیشتر قارهها فرصتی برای تنوع گونهای جدید دایناسورها خلق کرد.
کرتاسه
[ویرایش]کرتاسه طولانیترین دوره از دوران میانهزیستی (مزوزوئیک) را تشکیل میدهد اما شامل تنها دو دوره میشود: کرتاسه اولیه و پسین.[۱۷]

کرتاسه اولیه از ۱۴۵ تا ۱۰۰ میلیون سال پیش را پوشش میدهد.[۱۷] کرتاسه اولیه شاهد گسترش آبراهها و کاهش گونههای خزندهپایان، استگوسورها و دیگر گروههای گردنفراز[۱۸] با تمرکز کاهش سوسمارپایان در آمریکای شمالی بود.[۱۹] برخی دایناسورهای «جزیرهگَرد»، مثل «خوشخمیده»، برای وفق یافتن با کرانههای کمژرفا و جزایر کوچک اروپای باستان تکامل یافتند. دیگر دایناسورها از جمله کارکارودونتوسور و اسپینوسور برای پرکردن جای خالی که بهواسطه انقراض ژوراسیک-کرتاسه ایجاد شده بود، پدیدار شدند. بهتدریج شرایط فصلی دوباره پدیدار شده و مناطق قطبی در بعضی ایام سال سرد شد با اینحال بعضی دایناسورها، مثل لیالیناسورها و موتابوراسور همچنان طی تمام فصول، جنگلهای قطبی را بهمنزلگاه خود مبدل کرده بودند. مناطق قطبی بیش از حد معمول برای تمساحها سرد شده بود اما همچنان پناهگاه دوزیستان بزرگ مثل کولاسوکوس محسوب میشد. با تکامل سردههایی مانند کهنباشنده و پرندهدست، خزندگان بالدار (پتروسورها) بزرگتر شدند. همزمان دامنه پراکنش پستانداران وسیعتر میشد - هوسهقیفدندانسانان جانوران شکاری نسبتاً بزرگی مانند خزنده پستاندار و گوبیقیفدندان را تکامل دادند؛ از سویدیگر ددان نخستین شروع به شاخهشاخه شدن و در نتیجه تکامل پسددان و هوددان کردند؛ و گچیدندانهای[۲۰] چندتکمیزهای برای تداوم حضور در سنگوارهها، راه تکامل را درپیش گرفته بودند.
کرتاسه پسین از ۱۰۰ تا ۶۶ میلیون سال پیش گسترش دارد. کرتاسه پسین روند سرد شدن تدریجی هوا را تجربه کرد، روندی که در عصر نوزیستی نیز ادامه یافت. با گذشت زمان مناطق استوایی به خط استوا محدود شدند و مناطق خارج از مناطق استوایی، تغییرات فصلی شدیدی را در شرایط آب و هوایی متحمل شدند. دایناسورها همچنان شکوفا بودند و آرایههایی چون تیرانوسوروس، آنکیلوسور، تریسراتوپس و اردکمنقاران شبکه و زنجیره غذایی گیتی را تحت فرمان داشتند. در اقیانوسها موساسوریان که نقش ایکتیوسورها را پر کرده بودند، حکمرانی میکردند، موجوداتی که پس از افول و کاهش، در بین رویداد مرزی سنومانی-تورونی منقرض شدند. گرچه بیشخزندهواران نیز در رویداد مشابه منقرض شدند، گونههای گردندراز پلسیوسور مانند الاسموسور همچنان به رویش و تکثیر خویش ادامه میدادند. گیاهان گلدار، احتمالاً اولین بار طی دوره تریاس پدیدار شدند اما شکوفایی و غلبه واقعی را برای نخستین بار در کرتاسه تجربه کردند. خزندگان پرنده (پتروسورها) در دوره کرتاسه پسین بنا به دلایل چندان معلومی دچار افت شدند، اگرچه شاید این رخداد را بتوان در تمایلات سنگوارهها جستجو کرد و احتمالاً تنوع زیستی پتروسورها بسیار بالاتر از تصورات پیشین بودهاست. پرندگان بهطور فزایندهای رایجتر شده و به اشکال بسیار گوناگونی از ناهمسانمرغان و پرندهدنبان تکامل یافتند. اگرچه بهطور عمده کوچک ماندند ولی باخترمرغان دریایی نسبتاً بزرگ و فاقد قدرت پرواز گشتند چراکه به زندگی خود در دریاهای آزاد خو گرفته بودند. پسددان و هوددان نیز رایج گشتند و حتی گونههای بزرگ و تخصصیافته مثل دوزهداندندان[۲۱] و شوالتریا را پدیدآوردند. اما مهمترین پستانداران آن دوره همان چندتکمیزهایان بودند، مانند گچیدندانان در شمال و گندواناددسانان در جنوب.
در اواخر کرتاسه فعالیتهای آتشفشانی تلههای دکن[۲۲] و دیگر فورانها درحال آلوده کردن جَوّ بودند. با پدیدار شدن مستمر فعالیت آتشفشانی، این تصور قوت میگیرد که شهابی عظیمالجثه تقریباً ۶۶ میلیون سال پیش با زمین برخورد کرده و دهانه چیکشلوب را پدیدآورده است. (رویداد معروف به انقراض K-Pg یا کرتاسه – پالئوژن) سابقاً انقراض K-T، پنجمین و جدیدترین رویداد فجیع منقرض شدن جمعی است که طی آن تقریباً ۷۵٪ از تمام گونههای زیستی روی زمین از جمله تمام دایناسورهای غیر پرنده بهطور کامل از صفحهٔ گیتی زدوده شدند.[۲۳][۲۴][۲۵]
زمینساخت و جغرافیای دیرین
[ویرایش]در مقایسه با فرایند پرقدرت همگرایی صفحهای و کوهزایی در اواخر دیرینهزیستی (پالئوزوئیک)، فعالیت زمینساخت در دوران میانهزیستی (مزوزوئیک) نسبتاً ملایمتر بود. تنها کوهزایی بزرگ دوره مزوزوئیک در آنچه که حال حاضر شمالگان شناخته میشود، رخ داد. این رویداد منجر به ایجاد کوهزایی اینوئیتی،[۲۶] رشتهکوه بروکس، کوهستان ورخویانسک و رشتهکوه چرسکی در سیبری، و کوههای خینگان بزرگ در منچوری شد. این کوهزایی به گشایش اقیانوس منجمد شمالی و اتصال پوستههای پایای چین شمالی و سیبری به آسیا مربوط میشد.[۲۷] در مقابل، این دوران بهواسطه ازهمگسستگی عظیم ابرقارهٔ پانگهآ متبلور شد که بتدریج به یک قاره شمالی به نام لوراسیا و یک قاره جنوبی به نام گندوانا تقسیم گشت. این پویایی، حاشیههای غیرفعال قارهای پدیدآورد که امروزه ویژگی اکثر کرانههای اقیانوس اطلس (مانند سواحل شرقی ایالات متحده) محسوب میشوند.[۲۸]
تا پایان این دوره، قارهها تقریباً به شکل امروزی شان تفکیک شدند، هرچند همچنان موقعیتهایی متفاوت داشتند. لوراسیا به آمریکای شمالی و اوراسیا تبدیل شد، در حالی که گندوانا به آمریکای جنوبی، آفریقا، استرالیا، جنوبگان و شبهقارهٔ هند تقسیم شد. برخورد شبه قاره هند با صفحه آسیا طی دوره نوزیستی، به پیدایش رشتهکوه هیمالیا منجر شد.
آب و هوا
[ویرایش]دوره تریاس عموماً خشک بود؛ روندی که از اواخر کربنیفر آغاز شد و به خصوص در بخش داخله پانگهآ بهشدت شکل فصلی داشت. سطح پایین دریا نیز ممکن است منجر به تشدید دمای بیش از حد معمول میشدهاست. با وجود ظرفیت گرمایی ویژهٔ بالا، آب نقش یک منبع پایدارکننده در تنظیم حرارت محیط را دارد و نواحی خشکیِ نزدیک تودههای بزرگ مایع - بهخصوص اقیانوسها - نوساناتِ حرارتی کمتری تجربه میکنند. از آنجایی که بخش قابل توجهی از پانگهآ از کرانههای آن فاصله داشت، نوسانات دما شدید بود و احتمالاً نواحی داخلی دربرگیرنده بیابانهای پهناور بودند. لایههای قرمز فراوان و تبخیری مانند سنگ نمک، فرضیه فوق را پشتیبانی میکنند؛ اما شواهدی نیز موجود است که نشان میدهد خشکیِ فراگیر آب و هوایی دوره تریاس، بعضاً شاهد بارشهای فزاینده باران و دورههای با رطوبت بالا هم بودهاست.[۲۹] رویداد بارشزای کارنین مهمترین بازههای مرطوب محسوب میشد و دوره مرطوب دیگر در راتین، چندین میلیون سال پیش از رویداد انقراض مرزی تریاس-ژوراسیک، رخ داد.
سطوح آب دریاها در طول دوره ژوراسیک احتمالاً بر اثر گسترش بستر اقیانوس بالا آمد و افزایش یافت. شکلگیریِ پوسته جدید در زیر سطح زمین، تا مقدار ۲۰۰ متر (۶۵۶ فوت) بالاتر از سطح کنونی، آب اقیانوسها را بالا آورده و باعث جاری شدن سیل و محصور شدن نواحی ساحلی گشت. افزون بر این، پانگهآ شروع به تقسیم به تودههای کوچکتر کرد که طی آن خط ساحلی جدیدی پیرامون اقیانوس تتیس شکل گرفت. در این میانه، دما همزمان درحال افزایش بود که پس از مدتی شروع به ثابت شدن کرد. همینطور با توجه به افزایش نزدیکی نواحی خشکی و تودههای بزرگ آب، رطوبت نیز بهطور مداوم درحال افزایش بود که منجر به پسروی بیابانها شد.
تشخیص آب و هوای دوره کرتاسه از قطعیت کمتری برخوردار است و بیشتر مورد مناقشه و اختلاف نظر پژوهشگران میباشد. به احتمال فراوان سطوح بالاتری از دیاکسید کربن در جو زمین، دلیل و عامل حذف تقریبی شیب دمایی در راستای محور شمال به جنوب سیاره بودهاست. (دماها در سراسر سیاره تقریباً ثابت و ۱۰ درجه سانتیگراد گرمتر از دماهای فعلی بود). گردش اکسیژن در اعماق اقیانوسها نیز احتمالاً متوقف شده بود و این امر باعث تجزیه نشدن مقادیر عظیمی از مواد آلی شد که نهایتاً بهصورت نهشتهگذاریهای «سنگ شیل» بهجا ماندند.[۳۰][۳۱]
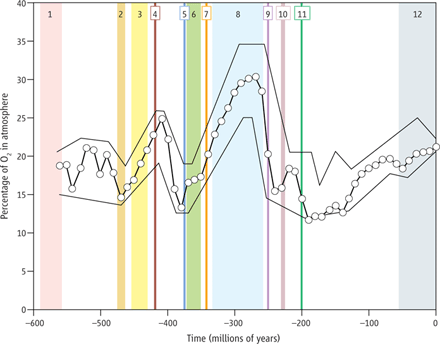
تحقیقات مختلف، منجر به نتایج جداگانهای از مقدار اکسیژن موجود در اتمسفر، در طول بخشهای مختلف میانهزیستی شدهاست: برخی مطالعات دلالت بر مقدار کمتر اکسیژن نسبت به سطح کنونی (۲۱٪) در طی تمام این دوره، دارند،[۳۲][۳۳] گروهی دیگر معتقدند کمبود اکسیژن در تریاس و بخشی از ژوراسیک کمتر بوده، ولی در کرتاسه میزان اکسیژن بالا بودهاست[۳۴][۳۵][۳۶] و گروهی دیگر سطح اکسیژن را در کل زمانهای تریاس، ژوراسیک و کرتاسه یا بخشی از آنها، بسیار بالاتر میدانند.[۳۷][۳۸]
زیست
[ویرایش]گیاهان
[ویرایش]
گونههای گیاهیِ غالبِ آن زمان بازدانگان بودند. اینها گیاهان آوندی مخروطدار هستند، که تخم بدون پوسته و پوشش را تولید میکنند. این حالت مغایر با ترکیب گیاهی فعلی در کره زمین است که گونههای غالب گیاهی را نهاندانگان (گیاهان گلدار) تشکیل میدهند. نخستین اعضای سردهٔ کهندار اولین بار در میانههای ژوراسیک پدیدار گشتند، این سرده امروز تنها بهواسطه یک گونه یعنی کهندار که تنها بازمانده این سرده بشمار میآید نمایندگی میشود.[۳۹] گروههای درختان مخروطیِ امروزی شروع به توسعه و ایجاد سردهها در ژوراسیک کردند.[۴۰] بنتیتسانان[۴۱] یکی از سردههای منقرض شده بازدانگان است. آنها دارای شاخ و برگهایی سطحی مشابه نخلیسرخسسانان بودند و در طول تریاس پسین، یکپارچه در دنیا پخش و منتشر شدند و از متداولترین خانوادههای گیاهی میانهزیستی بهشمار میآمدند.[۴۲]
گیاهان گلدار یا همان نهاندانگان اولین بار در اوایل دوره کرتاسه ظهور کردند، نخست در مناطق گرمسیری اما شیب یکنواخت دما به آنها اجازه گسترش به مناطق قطبی طی همه ادوار زمینشناسی را داد. در انتهای کرتاسه، نهاندانگان بر بیشتر مناطق درختی فرمانروایی میکردند هرچند که شواهد بیانگر توده زیستی غالب برای نخلیسرخسسانان و سرخسها (تا قبل از وقوع انقراض مرزی کرتاسه/پالئوژن) است. بعضی گونههای گیاهی انتشار متفاوت و ناملموسی از ادوار زمینشناسی بعد از خود داشتند. برای مثال، شانهسرخسسانان، دستهای از سرخسها هستند که در میانهزیستی در نیمکره شمالی پراکندگی خوبی داشتند، ولی اکنون بیشتر میتوان آنها را در نیمکره جنوبی یافت.[۴۳]
جانوران
[ویرایش]
انقراض تقریباً تمام گونههای جانوری در انتهای دوره پرمین شرایط را برای واگرایی سازشی بسیار از اشکال زندگیِ تازه مهیا کرد. بهطور خاص، انقراض گیاهخوار بزرگجثهای به نام پاریاسور و گوشتخواری به نام زشتسیما، کُنامهای زیستمحیطی را خالی گذاشتند. برخی ار این فضاهای تهی توسط سگدندانهای بازمانده و دوسگدندانها پر شد، که البته دومی متعاقباً خود منقرض شدهاست. پژوهشهای تازه حاکی از گذر زمانی دراز برای برقراری دوباره اکوسیستمهای پیچیده، با ساختار زیستیِ متنوع، شبکه غذایی درهم آمیخته و حیوانات تخصصی گوناگون در جایگاههای اکولوژیکی متفاوت، است. شروع بازسازی از بخش میانی تریاس یعنی ۴ تا ۶ میلیون سال پس از حادثه انقراض،[۴۴] تا شکوفایی و انتشار کامل ۳۰ میلیون سال بعد از رخداد انقراض به طول انجامید.[۴۵] بدینترتیب زندگی جانوری پس از انقراض پرمین-تریاس توسط شاهخزندگان (دایناسورها، پتروسورها و خزندگان آبی مانند ایکتیوسورها، پلسیوسورها و موساسوریان) تحت شعاع قرار گرفت.
تغییرات آب و هوایی اواخر ژوراسیک و کرتاسه موجب سازگاری و واگرایی هرچه بیشتر انواع گونههای زیستی شد. ژوراسیک اوج دوران گونهگونیِ شاهخزندگان، ظهور اولین پرندگان و پستانداران هودَد بود. شماری بر این اعتقاد هستند که حشرات همراه با نهاندانگان یا همان گیاهان گلدار بهعلت نوعی همزیستی رشد و نمو داشتهاند، زیرا کالبدشناسی آنها، بهخصوص بخشهای دهانی آنها برای تغذیه و همگامی با گیاهان گلدار مناسب و مساعد بهنظر میآید. هرچند تمام اجزاء اصلی فک و دهان حشرات به ظهور نهاندانگان منوط نمیشدند و تنوع گونهای جانوران حشرهای پس از پیدایش گیاهان گلدار حتی تقلیل پیدا کرد؛ بنابرین از منظر کالبدشناختی، کالبد حشرات از اهداف گوناگون دیگری نیز نشأت گرفته بودهاست.
ریزاندامگان
[ویرایش]در آغاز دوره میانهزیستی، جوامع پلانکتونهای اقیانوسی از نوعی که تحت تسلط آرکیپلاستیدانهای سبز بودند به نوعی دیگر که تحت تسلط جلبکهای همزیست با پلاستیدهای مشتقشده از جلبکهای قرمز بودند، تغییر پیدا کردند. گمان میرود که این دگرگونی بر اثر کمبود فزاینده بسیاری از فلزات کمیاب در دوره میانهزیستی رخ داده باشد.[۴۶]
جستارهای وابسته
[ویرایش]منابع
[ویرایش]- ↑ مُر، فرید؛ مدبری، سروش (۱۳۹۱). فرهنگ جامع علوم زمین. فرهنگ معاصر. شابک ۹۷۸-۶۰۰-۱۰۵-۰۴۰-۴.
- ↑ Bennettitales
- ↑ رجوع کنید به:
- Phillips, John (1840). [[۱](https://babel.hathitrust.org/cgi/pt?id=hvd.hn4zr7;view=1up;seq=163) "Palæozoic series"]. Penny Cyclopaedia of the Society for the Diffusion of Useful Knowledge. Vol. 17. London: Charles Knight and Co. pp. 153–54.
{{cite book}}: Check|chapter-url=value (help) "As many systems or combinations of organic forms as are clearly traceable in the stratified crust of the globe, so many corresponding terms (as Palæozoic, Mesozoic, Kainozoic, &c.) may be made, … " - Wilmarth, Mary Grace (1925). [[۲](https://books.google.com/books?id=my7x_PBkpm4C&pg=PA9) Bulletin 769: The Geologic Time Classification of the United States Geological Survey Compared With Other Classifications, accompanied by the original definitions of era, period and epoch terms]. Washington, D.C.: U.S. Government Printing Office. p. 9.
{{cite book}}: Check|url=value (help)
- Phillips, John (1840). [[۱](https://babel.hathitrust.org/cgi/pt?id=hvd.hn4zr7;view=1up;seq=163) "Palæozoic series"]. Penny Cyclopaedia of the Society for the Diffusion of Useful Knowledge. Vol. 17. London: Charles Knight and Co. pp. 153–54.
- ↑ الگو:Cite OEtymD
- ↑ Tang, Carol Marie. [[۳](https://www.britannica.com/science/Mesozoic-Era) "Mesozoic Era"]. Encyclopædia Britannica. Encyclopædia Britannica. Retrieved 5 September 2019.
{{cite web}}: Check|url=value (help) - ↑ Benton M J (2005). "Chapter 8: Life's Biggest Challenge". When life nearly died: the greatest mass extinction of all time. London: Thames & Hudson. ISBN 978-0-500-28573-2.[کدام صفحه؟]
- ↑ Stanley, Steven M. (3 October 2016). "Estimates of the magnitudes of major marine mass extinctions in earth history". Proceedings of the National Academy of Sciences of the United States of America. National Academy of Sciences. 113 (42): E6325–E6334. Bibcode:2016PNAS..113E6325S. doi:10.1073/pnas.1613094113. ISSN 1091-6490. PMC 5081622. PMID 27698119.
- ↑ Gradstein F, Ogg J, Smith A (2005). A Geologic Time Scale 2004. Cambridge University Press. ISBN 978-0-511-53604-5.
- ↑ Petersen, Sierra V.; Dutton, Andrea; Lohmann, Kyger C. (2016). "End-Cretaceous extinction in Antarctica linked to both Deccan volcanism and meteorite impact via climate change". Nature Communications. 7: 12079. Bibcode:2016NatCo...712079P. doi:10.1038/ncomms12079. PMC 4935969. PMID 27377632. Retrieved 31 July 2023.
- ↑ Alan Logan (31 January 2024). "Triassic Period". britannica.com.
- ↑ Ruta, Marcello; Benton, Michael J. (November 2008). "Calibrated Diversity, Tree Topology and the Mother of Mass Extinctions: The Lesson of Temnospondyls". Palaeontology. 51 (6): 1261–1288. doi:10.1111/j.1475-4983.2008.00808.x. S2CID 85411546.
- ↑ Rubidge. "Middle Triassic". palaeos.com.
- ↑ Capriolo, Manfredo; Marzoli, Andrea; Aradi, László E.; Callegaro, Sara; Corso, Jacopo Dal; Newton, Robert J.; Mills, Benjamin J. W.; Wignall, Paul B.; Bartoli, Omar; Baker, Don R.; Youbi, Nasrrddine; Remusat, Laurent; Spiess, Richard; Szabó, Csaba (7 April 2020). "Deep CO2 in the end-Triassic Central Atlantic Magmatic Province". Nature Communications. 11 (1): 1670. Bibcode:2020NatCo..11.1670C. doi:10.1038/s41467-020-15325-6. PMC 7138847. PMID 32265448. S2CID 215404768.
- ↑ Tegner, Christian; Marzoli, Andrea; McDonald, Iain; Youbi, Nasrrddine; Lindström, Sofie (26 February 2020). "Platinum-group elements link the end-Triassic mass extinction and the Central Atlantic Magmatic Province". Scientific Reports. 10 (1): 3482. doi:10.1038/s41598-020-60483-8. PMC 7044291. PMID 32103087. Retrieved 28 July 2023.
- ↑ ۱۵٫۰ ۱۵٫۱ Carol Marie Tang (7 February 2024). "Jurassic Era". britannica.com.
- ↑ Enchanted Learning. "Middle Jurassic". Enchanted Learning.
- ↑ ۱۷٫۰ ۱۷٫۱ Carl Fred Koch. "Cretaceous". britannica.com.
- ↑ (high-browsing groups)
- ↑ Butler, R. J.; Barrett, P. M.; Kenrick, P.; Penn, M. G. (March 2009). "Diversity patterns amongst herbivorous dinosaurs and plants during the Cretaceous: implications for hypotheses of dinosaur/angiosperm co-evolution". Journal of Evolutionary Biology. 22 (3): 446–459. doi:10.1111/j.1420-9101.2008.01680.x. PMID 19210589. S2CID 26000791.
- ↑ cimolodonts
- ↑ Didelphodon
- ↑ Deccan traps
- ↑ Becker, Luann (2002). "Repeated Blows". Scientific American. 286 (3): 76–83. Bibcode:2002SciAm.286c..76B. doi:10.1038/scientificamerican0302-76. PMID 11857903.
- ↑ "Cretaceous". University of California.
- ↑ Elizabeth Howell (3 February 2015). "K-T Extinction event". Universe Today.
- ↑ Innuitian orogeny
- ↑ Hughes, T. (August 1975). "The case for creation of the North Pacific Ocean during the Mesozoic Era". Palaeogeography, Palaeoclimatology, Palaeoecology. 18 (1): 1–43. Bibcode:1975PPP....18....1H. doi:10.1016/0031-0182(75)90015-2. Retrieved 21 July 2023.
- ↑ Stanley, Steven M. Earth System History. New York: W.H. Freeman and Company, 1999. شابک ۰−۷۱۶۷−۲۸۸۲−۶
- ↑ Preto, Nereo; Kustatscher, Evelyn; Wignall, Paul B. (15 April 2010). "Triassic climates – State of the art and perspectives". Palaeogeography, Palaeoclimatology, Palaeoecology. 290 (1–4): 1–10. Bibcode:2010PPP...290....1P. doi:10.1016/j.palaeo.2010.03.015. Retrieved 21 July 2023.
- ↑ Leckie, R. Mark; Bralower, Timothy J.; Cashman, Richard (September 2002). "Oceanic anoxic events and plankton evolution: Biotic response to tectonic forcing during the mid-Cretaceous: OCEANIC ANOXIC EVENTS AND PLANKTON EVOLUTION". Paleoceanography and Paleoclimatology. 17 (3): 13–1–13–29. doi:10.1029/2001PA000623.
- ↑ Turgeon, Steven C.; Creaser, Robert A. (17 July 2008). "Cretaceous oceanic anoxic event 2 triggered by a massive magmatic episode". Nature. 454 (7202): 323–326. Bibcode:2008Natur.454..323T. doi:10.1038/nature07076. PMID 18633415. S2CID 4315155.
- ↑ Robert A. Berner, John M. VandenBrooks and Peter D. Ward, 2007, Oxygen and Evolution. Science 27 April 2007, Vol. 316 no. 5824 pp. 557–58. A graph showing the reconstruction from this paper can be found here, from the webpage Paleoclimate – The History of Climate Change.
- ↑ Berner R. A. 2006 GEOCARBSULF: a combined model for Phanerozoic atmospheric O2 and CO2. Geochim. Cosmochim. Acta 70, 5653–64. See the dotted line in Fig. 1 of Atmospheric oxygen level and the evolution of insect body size by Jon F. Harrison, Alexander Kaiser and John M. VandenBrooks
- ↑ Berner, Robert A. , 2009, Phanerozoic atmospheric oxygen: New results using the GEOCARBSULF model. Am. J. Sci. 309 no. 7, 603–06. A graph showing the reconstructed levels in this paper can be found on p. 31 of the book Living Dinosaurs by Gareth Dyke and Gary Kaiser.
- ↑ Berner R. A. , Canfield D. E. 1989 A new model for atmospheric oxygen over phanerozoic time. Am. J. Sci. 289, 333–61. See the solid line in Fig. 1 of Atmospheric oxygen level and the evolution of insect body size by Jon F. Harrison, Alexander Kaiser and John M. VandenBrooks
- ↑ Berner, R, et al. , 2003, Phanerozoic atmospheric oxygen, Annu. Rev. Earth Planet. Sci. , V, 31, p. 105–34. See the graph near the bottom of the webpage Phanerozoic Eon بایگانیشده در ۲۷ آوریل ۲۰۱۳ توسط Wayback Machine
- ↑ Glasspool, I.J. , Scott, A.C. , 2010, Phanerozoic concentrations of atmospheric oxygen reconstructed from sedimentary charcoal, Nature Geosciences, 3, 627–30
- ↑ Bergman N. M. , Lenton T. M. , Watson A. J. 2004 COPSE: a new model of biogeochemical cycling over Phanaerozoic time. Am. J. Sci. 304, 397–437. See the dashed line in Fig. 1 of Atmospheric oxygen level and the evolution of insect body size by Jon F. Harrison, Alexander Kaiser and John M. VandenBrooks
- ↑ Balducci, Stan (2000). "Mesozoic Plants". fossilnews.com. Archived from the original on 23 January 2013. Retrieved 28 July 2023.
- ↑ Leslie, Andrew B.; Beaulieu, Jeremy; Holman, Garth; Campbell, Christopher S.; Mei, Wenbin; Raubeson, Linda R.; Mathews, Sarah (September 2018). "An overview of extant conifer evolution from the perspective of the fossil record". American Journal of Botany (به انگلیسی). 105 (9): 1531–1544. doi:10.1002/ajb2.1143. PMID 30157290. S2CID 52120430.
- ↑ Bennettitales
- ↑ Blomenkemper, Patrick; Bäumer, Robert; Backer, Malte; Abu Hamad, Abdalla; Wang, Jun; Kerp, Hans; Bomfleur, Benjamin (26 March 2021). "Bennettitalean Leaves From the Permian of Equatorial Pangea—The Early Radiation of an Iconic Mesozoic Gymnosperm Group". Frontiers in Earth Science. 9: 652699. Bibcode:2021FrEaS...9..162B. doi:10.3389/feart.2021.652699. ISSN 2296-6463.
- ↑ C.Michael Hogan. 2010. Fern. Encyclopedia of Earth. National council for Science and the Environment بایگانیشده در ۹ نوامبر ۲۰۱۱ توسط Wayback Machine. Washington, DC
- ↑ Lehrmann, D. J.; Ramezan, J.; Bowring, S.A.; et al. (December 2006). "Timing of recovery from the end-Permian extinction: Geochronologic and biostratigraphic constraints from south China". Geology. 34 (12): 1053–56. Bibcode:2006Geo....34.1053L. doi:10.1130/G22827A.1.
- ↑ Sahney, S. & Benton, M. J. (2008). "Recovery from the most profound mass extinction of all time". Proceedings of the Royal Society B: Biological Sciences. 275 (1636): 759–65. doi:10.1098/rspb.2007.1370. PMC 2596898. PMID 18198148.
- ↑ Zhang, Qiong; Bendif, El Mahdi; Zhou, Yu; Nevado, Bruno; Shafiee, Roxana; Rickaby, Rosalind E. M. (31 October 2022). "Declining metal availability in the Mesozoic seawater reflected in phytoplankton succession". Nature Geoscience. 15 (1): 932–941. doi:10.1038/s41561-022-01053-7. hdl:10451/55860. Retrieved 21 July 2023.
{kind=link}