
گونهزایی بومشناختی

گونهزایی بومشناختی (به انگلیسی: Ecological speciation)، شکلی از گونهزایی ناشی از جدایی تولیدمثلی است که به دلیل یک عامل بومشناختی که جریان ژن را بین دو جمعیت از یک گونه کاهش میدهد یا حذف میکند، رخ میدهد. عوامل بومشناختی میتواند شامل تغییر در شرایط محیطی باشد که یک گونه در آن تجربه میکند، مانند تغییرهای رفتاری شامل شکار، اجتناب از شکارچی، جذب گردهافشان و جستجوی غذا. و همچنین تغییر در گزینش جفت به دلیل گزینش جنسی یا سیستمهای ارتباطی. جدایی تولیدمثلی مبتنی بر محیط زیست تحت فرگشت واگرای متفاوت سبب تشکیل گونههای جدید میشود. این پدیده در بسیاری از موارد در طبیعت ثبت شدهاست و در چند دهه گذشته تمرکز اصلی پژوهشها دربارهٔ گونهزایی بودهاست.[۱] : 179
| نوع جداسازی باروری | پیش از تخمک (زیگوتیک) یا پس از تخمک (زیگوتیک) | علت بومشناختی انتخاب | |||
|---|---|---|---|---|---|
| محیطهای واگرا | برهمکنشهای بومشناختی | گزینش جنسی | تقویت | ||
| زیستگاه | پیش | ✓ | ✓ | ✓ | |
| جنسی/گردهافشان | پیش | ✓ | ✓ | ✓ | ✓ |
| زمانی | پیش | ✓ | ✓ | ✓ | |
| انتخاب در برابر مهاجران | پیش | ✓ | ✓ | ✓ | |
| پس از جفتگیری | پیش | ✓ | ✓ | ✓ | ✓ |
| انتخاب در برابر دورگهها | پس | ✓ | ✓ | ✓ | ✓ |
| مستقل از محیط زیست | پس | ✓ | ✓ | ✓ | ✓ |
| وابسته به محیط زیست | پس | ✓ | ✓ | ||
انواع جدایی تولیدمثلی[ویرایش]
- جدایی زیستگاه
- جغرافیایی
- جدایی جنسی
- جدایی گردهافشان
- جدایی گردهافشان مکانیکی
- جدایی گردهافشان رفتاری
- جدایی زمانی (گونهزایی ناهمزمانی)
- دیگر شکلهای جدایی بومشناختی پیش از تخمک
- شکلهای جدایی بومشناختی پس از تخمک
_Schematic.svg/310px-Ecological_Speciation_(habitat_isolation)_Schematic.svg.png)




| گونهها | ویژگی |
|---|---|
| Acropora spp. | مرجانهای ژاپنی با زمان تخمریزی خود از نظر تولیدمثلی جدا میشوند.[۳] |
| Montastraea annularis , M. faveolata و M. franksi | سه گونه مرجانی مرتبط که به دلیل زمان تخمریزی خود گونهزایی کردهاند.[۴] |
| Oncorhynchus nerka | دورههای پرورش سالانه ماهی سالمون Sockeye در دو دوره در سال (اواخر و اوایل) رخ میدهد و باعث جدایی ژنتیک جمعیتهای اولیه شدهاست. پرورش ماهی قزلآلا به عنوان ژنتیک شناخته شدهاست اما هیچ ژن خاصی برای این گونه شناخته شده نیست.[۵][۶][۷] |
| Thaumetopoea pityocampa | تسلط همزمان در ژنها با زمان ظهور مراحل لاروی این گونه پروانه همراه است. جمعیت لاروهای زمستانی و تابستانی در حال گونهزایی هستند.[۸][۹][۱۰] |
| Inurois punctigera | در مناطقی که دمای اواسط زمستان برای گونه شبپره نامناسب است از پرورش جلوگیری میشود. این امر باعث ایجاد جمعیتهای پسین و پیشین شدهاست.[۱۱] |
| Pemphigus populi-transversus و P. obesinymphae | شتههای گالی در برگهای مختلف گونههای درخت میزبان یکسان تولید میکنند. P. populi-transversus روی برگهای اوایل بهار گال ایجاد میکند، در حالی که P. obesinymphae آنها را روی برگها تابستانه تشکیل میدهد. این سبب انزوای کامل باروری شدهاست.[۱۲] |
| Asphondilia spp. | سه گونه میج ساقههای Larrea tridentata , A. auripila در تابستان، A. resinosa در زمستان و A. foliosa در بهار را آلوده میکنند.[۱۳] |
| Acropora samoensis | جمعیت گونههای همجا مرجانها بهطور جداگانه در پاییز و بهار تخمریزی میکنند و تخمریزی یک امر ارثی است که احتمالاً شامل ژن PaxC میشود.[۱۴] |
| Cellana spp. | در ژرفای مختلف در عرض چند سانتیمتر، خارهچسبها به دلیل ترکیبی از گونهزایی و نشانههای تخمریزی (مانند تخمریزی بر اساس سطح آب) از نظر تولیدمثلی جدا شدهاند.[۱۵] |
| Hydrobates spp. | گروه پترلها از نظر تولیدمثلی گونههای جداشده (در آزورها) و گونههای آغازین (دیگر مجمعالجزایر) ناشی از فصلهای تولیدمثلی سرد و گرم را دارند.[۱۶][۱۷][۱۸] |
| Howea belmoreana و H. forsteriana | زمان گلدهی کنترلشده ژنتیکی باعث جدایی تولیدمثلی دو گونه نخل در جزیره لرد هاو (در ارتباط با سطح مختلف pH خاک) شدهاست.[۱۹] |
| قارچ سفیدک سطحی انگور | شواهدی از جدایی به دلیل تفاوتهای زمانی گونه میزبان آن Vitis vinifera را نشان میدهد.[۲۰] |
| Oncorhynchus gorbuscha | چرخههای زندگی دو ساله زوج و فرد در ارتباط با دورههای پرورش فصلی ماهی قزلآلای صورتی باعث تمایز ژنتیکی بین این دو جمعیت شدهاست.[۲۱][۲۲][۲۳] |
| Magicicada spp. | گروههایی متشکل از جفتهای چرخه زندگی ۱۳ و ۱۷ ساله (در مجموع هفت گونه) از زنجرهها ظاهر میشوند تا با بازههای زمانی بزرگ بین فصلهای نازایی از هم جدا شوند.[۲۴][۲۵][۲۶] فقط هر ۲۲۱ سال، چرخههای ۱۳ و ۱۷ ساله در جایی که هر دو جفت بهطور همزمان ظاهر میشوند، هماهنگ میشوند.[۲۷] |
| Antitrogus parvulus | دو گروه سوسک تمایز ژنتیکی را از چرخههای زندگی که با فاصلۀ دو ساله جدا میشوند، بیان میکنند.[۲۸] |
| Oeneis melissa semidea | چرخه زندگی دو ساله گروههای زادآور گونههای پروانه باعث تمایز ژنتیکی شدهاست.[۲۹] |
| Bambusoideae | بامبوها در حال تولیدمثل سمبلزا هستند، جایی که سالها قبل از گلدهی انبوه در آن زندگی میکنند. این میتواند در سالهای مختلف و مکانهای مختلف رخ دهد. گمان میرود که لکههای آلوکرونیک باعث تنوع گونههای جهانی بامبو شدهاند.[۳۰][۳۱][۳۲] |
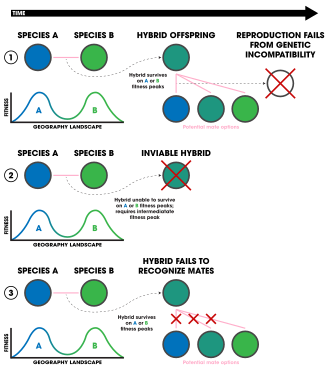
1. جدایی پس از زیگوت مستقل از محیط زیست.
2. جدایی پس از زیگوت وابسته به محیط زیست.
3. انتخاب در برابر دورگهها
منابع[ویرایش]
- ↑ Jerry A. Coyne; H. Allen Orr (2004), Speciation, Sinauer Associates, pp. 1–545, ISBN 0-87893-091-4
- ↑ Howard D. Rundle & Patrik Nosil (2005), "Ecological speciation", Ecology Letters, 8 (3): 336–352, doi:10.1111/j.1461-0248.2004.00715.x
- ↑ H. Fukami, M. Omori, K. Shimoike, T. Hayashibara, and M. Hatta (2003), "Ecological and genetic aspects of reproductive isolation by different spawning times in Acropora corals", Marine Biology, 142 (4): 679–684, doi:10.1007/s00227-002-1001-8
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ N. Knowlton, J. L. Maté, H. M. Guzmán, R. Rowan, and J. Jara (1997), "Direct evidence for reproductive isolation among the three species of the Montastraea annularis complex in Central America (Panamá and Honduras)", Marine Biology, 127 (4): 705–711, doi:10.1007/s002270050061
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Andrew P. Hendry, Ole K. Berg, and Thomas P. Quinn (1999), "Condition dependence and adaptation-by-time: breeding date, life history, and energy allocation within a population of salmon", Oikos, 85 (3): 499–514, doi:10.2307/3546699, JSTOR 3546699
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Andrew P Hendry, Yolanda E Morbey, Ole K Berg, and John K Wenburg (2004), "Adaptive variation in senescence: reproductive lifespan in a wild salmon population", Proceedings of the Royal Society B: Biological Sciences, 271 (1536): 259–266, doi:10.1098/rspb.2003.2600, PMC 1691593, PMID 15058436
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ E. K. Fillatre, P. Etherton, and D. D. Heath (2003), "Bimodal run distribution in a northern population of sockeye salmon (Oncorhynchus nerka): life history and genetic analysis on a temporal scale", Molecular Ecology, 12 (7): 1793–1805, doi:10.1046/j.1365-294x.2003.01869.x, PMID 12803632
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ C.Pimentel, T.Calvão, M.Santos, C.Ferreira, M.Neves, and J. -Å.Nilsson (2006), "Establishment and expansion of a Thaumetopoea pityocampa (Den. & Schiff.) (Lep. Notodontidae) population with a shifted life cycle in a production pine forest, Central-Coastal Portugal", Forest Ecology and Management, 233 (1): 108–115, doi:10.1016/j.foreco.2006.06.005
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Helena M Santos, Maria-Rosa Paiva, Susana Rocha, Carole Kerdelhué, and Manuela Branco (2013), "Phenotypic divergence in reproductive traits of a moth population experiencing a phenological shift", Ecology and Evolution, 3 (15): 5098–5108, doi:10.1002/ece3.865, PMC 3892371, PMID 24455139
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Manuela Branco, Maria-Rosa Paiva, Helena Maria Santos, Christian Burban, and Carole Kerdelhué (2017), "Experimental evidence for heritable reproductive time in 2 allochronic populations of pine processionary moth", Insect Science, 24 (2): 325–335, doi:10.1111/1744-7917.12287, PMID 26530538
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Satoshi Yamamoto and Teiji Sota (2012), "Parallel allochronic divergence in a winter moth due to disruption of reproductive period by winter harshness", Molecular Ecology, 21 (1): 174–183, doi:10.1111/j.1365-294X.2011.05371.x, PMID 22098106
- ↑ Patrick Abbot and James H Withgott (2004), "Phylogenetic and molecular evidence for allochronic speciation in gall-forming aphids (Pemphigus)", Evolution, 58 (3): 539–553, doi:10.1111/j.0014-3820.2004.tb01677.x, PMID 15119438
- ↑ Jeffrey B Joy and Bernard J Crespi (2007), "Adaptive radiation of gall-inducing insects within a single host-plant species", Evolution, 61 (4): 784–795, doi:10.1111/j.1558-5646.2007.00069.x, PMID 17439611
- ↑ Natalie L Rosser (2015), "Asynchronous spawning in sympatric populations of a hard coral reveals cryptic species and ancient genetic lineages", Molecular Ecology, 24 (19): 5006–5019, doi:10.1111/mec.13372, PMID 26339867
- ↑ Christopher E. Bird, Brenden S. Holland, Brian W Bowen, and Robert J Toonen (2011), "Diversification of sympatric broadcast-spawning limpets (Cellana spp.) within the Hawaiian archipelago", Molecular Ecology, 20 (10): 2128–2141, doi:10.1111/j.1365-294X.2011.05081.x, PMID 21481050
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ L. R. Monteiro (1998), "Speciation through temporal segregation of Madeiran storm petrel (Oceanodroma castro) populations in the Azores?", Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 353 (1371): 945–953, doi:10.1098/rstb.1998.0259, PMC 1692297
- ↑ V. L. Friesen, A. L. Smith, E. Gómez-Díaz, M. Bolton, R. W. Furness, J. González-Solís, and L. R. Monteiro (2007), "Sympatric speciation by allochrony in a seabird" (PDF), PNAS, 104 (47): 18589–18594, Bibcode:2007PNAS..10418589F, doi:10.1073/pnas.0700446104, PMC 2141821, PMID 18006662
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Mark Bolton, Andrea L. Smith, Elena Gómez‐díaz, Vicki L. Friesen, Renata Medeiros, Joël Bried, Jose L. Roscales, and Robert W. Furness (2008), "Monteiro's Storm‐petrel Oceanodroma monteiroi: a new species from the Azores", Ibis, 150 (4): 717–727, doi:10.1111/j.1474-919X.2008.00854.x
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Vincent Savolainen, Marie-Charlotte Anstett, Christian Lexer, Ian Hutton, James J Clarkson, Maria V Norup, Martyn P Powell, David Springate, Nicolas Salamin, and William J Baker (2006), "Sympatric speciation in palms on an oceanic island", Nature, 441 (7090): 210–213, Bibcode:2006Natur.441..210S, doi:10.1038/nature04566, PMID 16467788
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Josselin Montarry, Philippe Cartolaro, Sylvie Richard-Cervera, and François Delmotte (2009), "Spatio-temporal distribution of Erysiphe necator genetic groups and their relationship with disease levels in vineyards", European Journal of Plant Pathology, 123: 61–70, doi:10.1007/s10658-008-9343-9
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Lev A. Zhivotovsky, A. J. Gharrett, A. J. McGregor, M. K. Glubokovsky, and Marcus W. Feldman (1994), "Gene differentiation in Pacific salmon (Oncorhynchus Sp.): facts and models with reference to pink salmon (O. Gorbuscha)", Canadian Journal of Fisheries and Aquatic Sciences, 51: 223–232, doi:10.1139/f94-308
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ D. Churikov and A. J. Gharrett (2002), "Comparative phylogeography of the two pink salmon broodlines: an analysis based on a mitochondrial DNA genealogy", Molecular Ecology, 11 (6): 1077–1101, doi:10.1046/j.1365-294x.2002.01506.x, PMID 12030984
- ↑ Morten T. Limborg, Ryan K. Waples, James E. Seeb, and Lisa W. Seeb (2014), "Temporally Isolated Lineages of Pink Salmon Reveal Unique Signatures of Selection on Distinct Pools of Standing Genetic Variation", Journal of Heredity, 105 (6): 835–845, doi:10.1093/jhered/esu063, PMID 25292170
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ D. C. Marshall and J. R. Cooley (2000), "Reproductive character displacement and speciation in periodical cicadas, with description of new species, 13-year Magicicada neotredecem", Evolution, 54 (4): 1313–1325, doi:10.1111/j.0014-3820.2000.tb00564.x, PMID 11005298
- ↑ C. Simon, J. Tang, S. Dalwadi, G. Staley, J. Deniega, and T. R. Unnasch (2000), "Genetic evidence for assortative mating between 13-year cicadas and sympatric "17-year cicadas with 13-year life cycles" provides support for allochronic speciation", Evolution, 54 (4): 1326–1336, doi:10.1111/j.0014-3820.2000.tb00565.x, PMID 11005299
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Teiji Sota, Satoshi Yamamoto, John R. Cooley, Kathy B. R. Hill, Chris Simon, and Jin Yoshimura (2013), "Independent divergence of 13- and 17-y life cycles among three periodical cicada lineages", PNAS, 110 (17): 6919–6924, Bibcode:2013PNAS..110.6919S, doi:10.1073/pnas.1220060110, PMC 3637745, PMID 23509294
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Rebecca S. Taylor and Vicki L. Friesen (2017), "The role of allochrony in speciation", Molecular Ecology, 26 (13): 3330–3342, doi:10.1111/mec.14126, PMID 28370658
- ↑ D. P. Logan, P. G. Allsopp, and M. P. Zalucki (2003), "Overwintering, soil distribution and phenology of Childers canegrub, Antitrogus parvulus (Coleoptera: Scarabaeidae) in Queensland sugarcane", Bulletin of Entomological Research, 93 (4): 307–314, doi:10.1079/ber2003245, PMID 12908916
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ A. E. Gradish, N. Keyghobadi, and G. W. Otis (2015), "Population genetic structure and genetic diversity of the threatened White Mountain arctic butterfly (Oeneis melissa semidea)", Conservation Genetics, 16 (5): 1253–1264, doi:10.1007/s10592-015-0736-y
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Madhav Gadgil and S. Narendra Prasad (1984), "Ecological Determinants of Life History Evolution of Two Indian Bamboo Species", Biotropica, 16 (3): 161–172, doi:10.2307/2388050, JSTOR 2388050
- ↑ Donald C. Franklin (2004), "Synchrony and asynchrony: observations and hypotheses for the flowering wave in a long‐lived semelparous bamboo", Journal of Biogeography, 31 (5): 773–786, doi:10.1111/j.1365-2699.2003.01057.x
- ↑ Anelena L. de Carvalho, Bruce W. Nelson, Milton C. Bianchini, Daniela Plagnol, Tatiana M. Kuplich, and Douglas C. Daly (2013), "Bamboo-Dominated Forests of the Southwest Amazon: Detection, Spatial Extent, Life Cycle Length and Flowering Waves", PLOS ONE, 8 (1): e54852, Bibcode:2013PLoSO...854852C, doi:10.1371/journal.pone.0054852, PMC 3554598, PMID 23359438
{{citation}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link)